
Objectives
Yukon River Chinook salmon (Oncorhynchus tshawytscha) are an important subsistence, commercial, and recreational resource. With runs once numbering close to 300,000 adults per year, these fatty fish were once the mainstay of subsistence communities and commercial fisheries all along the river. However, since the late 1990s, Chinook salmon returns have been in a period of prolonged decline with far fewer fish returning to the river to spawn. To help restore stocks, managers implemented measures to control Chinook salmon harvest. Commercial and sport fisheries were closed and subsistence fishing has been severely restricted in recent years. As a result, many of the hundreds of fish camps that once lined the Yukon River have disappeared or been abandoned. These restrictions have severely impacted and continue to impact communities located from the river mouth to the headwaters in Canada by eliminating access to the Chinook salmon resource that people have traditionally and historically relied on for their lives and livelihoods.
The causes of the Yukon River Chinook population declines and fluctuations are not known, but there is strong evidence that the early marine period, when the salmon transition from freshwater to the marine environment, represents a critical stage in juvenile salmon survival. Yukon River Chinook salmon are categorized as “stream-type Chinook” which spend one or two winters in freshwater before migrating to estuaries and eventually to the sea. During the early marine period, the fish must feed and grow to obtain sufficient size and store energy in preparation for their first winter at sea. There is evidence that the juvenile salmon need to reach a minimum size by the end of their first summer at sea in order to have good probability of avoiding predation and surviving the winter. This size-selective mortality suggests that the way the juvenile fish use their habitat, the abundance and quality of food, and the number of predators may all be very important in determining whether the fish grow to become adults to return to the river and spawn.
Another factor potentially affecting juvenile salmon survival is the match/mismatch between outmigration timing and available prey resources in the early marine life. Statistical models of outmigrating juvenile Chinook salmon dynamics have found a correlation between the timing of ice breakup at Dawson, Yukon Territories, in the year that smolts migrate to sea and the peak migration of fish leaving the river. If prey abundance matches the timing of outmigration, the juvenile fish will be larger and healthier when they reach the marine environment. Understanding the interplay of growth, outmigration timing, size, diet, and condition of juvenile Chinook salmon throughout early marine rearing habitats is, therefore, integral to addressing the role of early marine processes in structuring cohort strength of Yukon River stocks.
Methods

Sampling stations on the lower Yukon River and pro-delta. Red=south tributary, yellow = middle tributary, blue = north tributary, lines = pro-delta transects.
The Yukon River flows through the Yukon-Kuskokwim Delta, a large alluvial plain of sediment deposited by the Yukon and Kuskokwim Rivers over long periods of time. At its lower end, the Yukon River branches into three main tributaries (or mouths), each with numerous minor tributaries, sloughs, and channels. We partnered with local fishermen from the villages of Emmonak and Alakanuk to help us establish and sample three locations on each of the three mouths. Although the Yukon River can be several miles wide, in many places it is extremely shallow. The river is constantly changing, and there are numerous shallow mid-channel bars that cannot easily be seen in the silty waters. Local knowledge of the river has been critical for success of this research.
In collaboration with the Alaska Department of Fish and Game and the Yukon Delta Fisheries Development Association, and with financial support from the Arctic-Yukon-Kuskokwim Sustainable Salmon Imitative and the North Pacific Research Board, NOAA Fisheries scientists at the Alaska Fisheries Science Center initiated a study in 2014 to characterize habitat usage, size, diet, marine entry timing, and condition of juvenile Chinook salmon in the Yukon River Delta. Prior to this research, the only information on lower Yukon River Chinook salmon came from a single study of the southern tributary conducted in 1986. By combining net sampling methods in nearshore (tributary and tidal channel) and offshore (delta front and pro-delta) locations with oceanographic measurements, we hope to shed light on the complex processes affecting early juvenile Chinook salmon survival.

Setting the CTD instrument onboard the chartered fishing vessel Anchor Point.
At each of the sampling locations (or stations), we used a net towed between two outboard-powered skiffs to catch juvenile salmon and other fish. For each tow, we also collected information on water temperature, depth, and how fast the water was flowing through our nets. We sampled each station three times per week from the middle of May (just after the ice had left the river) to the end of July in 2014 and 2015.
As the Yukon River water passes the mouths of the tributaries, it spreads out across a subtidal area called the platform. The platform is large and extremely shallow, averaging less than 2 meters (6 feet) deep and extending as much as 30 kilometers (18 miles) off shore of the river mouths. Within the platform are a number of channels that have been scoured out by ice and river flow. Juvenile salmon must cross this platform as they head out to sea, but most of the area is too shallow to sample using trawls, and the water is too turbid to be able to locate the ice-scour channels that the fish may be using on their outmigration.
At its seaward edge, the platform drops off abruptly into deeper water in an area referred to as the delta front or pro-delta. The plume of freshwater from the Yukon River reaches all the way out to the pro-delta where it mixes with the marine waters of the northern Bering Sea. This is the seaward edge of the Yukon River estuary. We chartered the 42-foot fishing vessel Anchor Point out of Nome, Alaska, to sample five transects set perpendicular to the delta front (Fig. 1). These transects cross the river plume, the location of which changes seasonally throughout the sampling period, allowing us to intercept the fish as they begin the marine phase of their life.
On each transect, sampling stations are located at the 7-m, 10-m and 14-m water depth contours. Surveys conducted by AFSC scientists in the Bering Sea regularly sample as shallow as 20 m near the Yukon Delta. The depths of our stations were chosen to extend sampling inshore to the edge of the pro-delta. At each of our sampling stations, we used a surface trawl called a Mamou to catch salmon and other fish in the top 2 m of the water column. At the 10-m and 14-m stations, we also used a midwater Aluette trawl to sample the water column between 2-m and 5-m deep. Together, these nets can not only help us fish the top 5 m of the water column, but also can help us determine whether the fish we catch are on the surface of the water or deeper down.
A CTD (conductivity-temperature-depth) instrument fitted with a turbidity meter was used to collect information on the physical environment at each mile along the transects. After processing the information on a computer, we can see vertical profiles of the water that show how the temperature, salinity, and turbidity of the water vary as we move from shallow stations to deeper stations, as we move around the delta, and as we sample over the summer. Delta-front fish sampling and oceanographic data collection occurred on cruises in June, July, and August of 2014 and 2015.
All of the fish and invertebrates that we captured, both in the tributaries and off the pro-delta, were identified to species and counted. A subsample of each species and/or age group was measured, and any species we could not identify in the field was frozen or preserved in formaldehyde to be brought back to the Center’s Auke Bay Laboratory in Juneau for identification. Because this is only the second survey to be conducted in this area since 1986, we also collected whole fish vouchers of each species for inclusion in the University of Alaska Fairbanks museum.
Chinook and coho salmon captured in the river or on the pro-delta were measured, bagged, and either frozen or placed in formaldehyde so that they could be brought back to the lab to be analyzed. The energetic status of a subsample of juvenile Chinook and coho was analyzed to understand the condition of the fish. Prey resources of juvenile Chinook and coho were evaluated by removing the stomachs, sorting the stomach contents by prey type, and weighing each component.
Discussion

Figure 2. Catch per unit effort of juvenile Chinook salmon in the southern tributary by year and week (CPUE expressed as fish/10 minute tow).
To understand how changes in adult Chinook salmon runs have affected juvenile Chinook salmon abundance and distribution, we compared data from the first 2 years of the present study with the only prior study of lower Yukon River juvenile Chinook salmon which was undertaken in 1986 when salmon were abundant and returns were high. In 1986, tributary sampling was restricted to stations in the south tributary. Two of the sampling stations used in 2014 and 2015 were located as close as possible to stations sampled in 1986 to further a comparison between these studies. Using data from these stations, we found that catch per unit effort (CPUE) and outmigration timing were highly variable between the 2 years of our study and when compared with 1986 (Fig. 2). The CPUE was generally higher in 1986 over a longer period of time than in 2014 and 2015, which may reflect the higher abundance of salmon in the river, but we do not have enough data at present to speculate on the much lower CPUE in 2014 as compared with 2015. In the southern tributary, outmigration patterns for 2015 and 1986 are similar, with two distinct peaks of outmigration occurring approximately 2 weeks apart. This pattern is much less evident in 2014.


Figure 3. Comparison of CPUE between the three main Lower Yukon tributaries in 2014 and 2015 (CPUE expressed as fish/cubic meter of water).
A primary contribution of this research was to expand the spatial scale of sampling to include all three of the main lower tributaries, and additionally sample 15 stations on the pro-delta between the south tributary and Stuart Island east of the north tributary mouth. Comparing CPUE from 2014 and 2015 across tributaries highlights spatial and temporal differences in juvenile Chinook salmon habitat use and outmigration strategies (Fig. 3). For 2014, the dual peaks in outmigration that were absent from the south tributary (SM) data become more evident in the northern (NM) and middle (MM) tributary data. For 2015, all tributaries show more than one peak in outmigration, with the timing of these peaks occurring at different times on the southern tributary than on the northern and middle tributaries. These peaks may represent different cohorts of fish or may indicate that fish from different parts of the Yukon River system have distinct migration corridors in the individual tributaries. We hope that genetic samples collected from all of the juvenile Chinook salmon that were captured can help us tease out whether spatial and temporal differences in migration patterns represent distinct groups of fish.
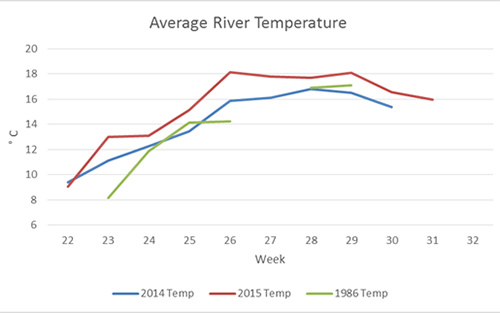
Figure 4. Comparison of average water temperatures between years.
Physical factors may also influence outmigration timing. Average water temperatures were approximately 1.3°C higher in 2015 than in either 2014 or 1986, and the water temperature increased faster and stayed higher longer (Fig. 4). For the southern tributary stations, the maximum water temperature of approximately 17 °C occurred near the second week of July in 1986 and 2015. In 2015, water temperatures reached a high of 18.1°C on June 26 and remained above 18°C until the July 17. In 1986 and 2015 the first outmigration peak in the southern tributary occurred when water temperatures were close to 14 °C, an event that happened two weeks earlier in 2015 than in 1986. When the average length of the fish is compared across the3 years (Fig. 5), it is evident that fish in the first peak of outmigration in 2015 were substantially smaller than fish from the same period in 2014 and 1986. This may suggest that temperature induced fish to migrate from the river earlier in 2015 and before they had the opportunity to feed and grow.
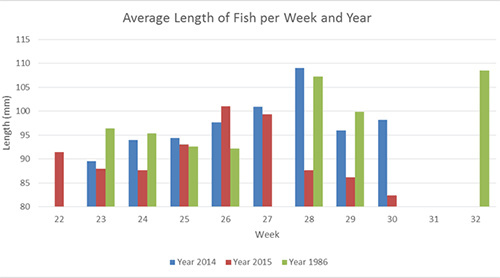
Figure 5. Juvenile Chinook salmon CPUE by tributary in 2015.
Prolonged high temperatures in 2015 may also have affected the type and availability of prey. Fish caught near the end of the sampling period in both 2014 and 1986 are significantly larger than fish from 2015 during the same timeframe. Limited data from stomach content analyses for 2014 indicates that fish are primarily consuming terrestrial drift and the larval stages of aquatic insects. A comparison of 2014 and 2015 stomach contents may highlight differences in prey type and stomach fullness. Spatial and temporal differences in stomach contents and stomach fullness may also occur across tributaries. To further investigate the role of prey types and availability on fish size and condition, our 2016 field data collection will include sampling for prey items, such as invertebrate drift in the tributaries. We will be assessing not only what prey items are available for the fish to eat, but also the nutritional quality of the prey they consume.
Results

Catch from the two-boat tow net. Photo NOAA Fisheries.
A second component of this research involved sampling along five transects placed perpendicular to the pro-delta. These transects cross the Yukon River plume front and provide an opportunity to investigate juvenile Chinook salmon in an estuarine environment. On the pro-delta, the majority of juvenile Chinook salmon were captured at the three northernmost stations with only a small proportion of the catch occurring at the station directly off the southern tributary mouth (SM). This was a surprising finding given the high CPUE of sampling in the southern tributary. As the Yukon river flows outward toward the ocean, the earth’s rotation causes it to be deflected northward, a phenomenon known as the Coriolis effect. This induces a general north-northeasterly flow of water around the delta. The higher CPUE for juvenile Chinook in the northern stations suggests that the fish may be following these prevailing currents north into Norton Sound possibly to access more abundant food resources or to escape predation.
Species composition between pro-delta transects and between transect stations varies seasonally, possibly in relation to the strength and location of the river plume. Nearly all the salmon were captured in the upper 3 meters of the water column, and abundance was higher in areas of active mixing between the river and marine waters. Post-sampling processing of CTD and catch data suggests that juvenile Chinook salmon may be utilizing these frontal boundary areas either as a transitional environment or for foraging. At the present time, we do not have the equipment to detect these fronts in the field during the sampling. Occasionally, we can detect a color change between the river and marine waters, and we have conducted opportunistic sampling in these areas. However, prevailing weather conditions on the pro-delta do not make this a reliable or consistent method of front detection. We have investigated the use of an airborne drone equipped with thermal and visual cameras that could detect the location of these frontal boundaries and assist with targeted sampling of these areas. However, at the present time we have not obtained funding to pursue this method of data collection.
Catch of juvenile Chinook salmon on the pro-delta was much lower than catch in the river, with the majority of fish captured in June. Juvenile Chinook salmon are captured during surveys in Norton Sound and the southern Bering Sea in late August and early September, suggesting that the salmon may have a relatively short period of residency in the Yukon River plume before heading fully into marine waters. This would be similar to findings of juvenile Chinook salmon use of plume environments in other large river systems.
Although the focus of this research was on juvenile Chinook salmon, we have also collected a large amount of data on other juvenile and adult fish in both the tributary and pro-delta habitats. Juvenile coregonids (whitefish and ciscos), burbot, and shellfish were the most abundant species in our tributary sampling in 2014 and 2015. Arctic lamprey, both the larval ammocoete and smolt stages, were also regular components of our tributary catch, and the smolt stage was also captured on the pro-delta. In 2016 we will be providing genetic samples to the University of Alaska to further research on this unusual species.
On the pro-delta, the location of the seaward edge of the Yukon River plume changes throughout the summer. In June when river discharge peaks, the warmer river water extends as a wedge across the entire platform forming a distinct front with the offshore marine waters. Later in the summer, this structure breaks down and, depending on the level of discharge, the river water spreads out in a layer that can extend to the outermost sampling stations. Temporal and spatial changes in stratification and temperature affect community composition on the pro-delta between sampling stations on a transect and between transect locations. The most abundant species in the pro-delta samples are juvenile and larval Pacific herring, juvenile and larval rainbow smelt, juvenile saffron cod, and nine-spine sticklebacks.
Conclusion
We have provided an overview of ongoing research into juvenile Chinook salmon habitat use, out-migration, and condition on the Yukon Delta. Preliminary data analysis shows interannual variation in juvenile Chinook salmon abundance and out-migration timing. CPUE in the 2 years of this study was lower than in 1986 when Chinook salmon abundance was high, but migration patterns between the study years are similar. Our research shows spatial and temporal differences in outmigration timing and strength between the three main Lower Yukon tributaries, which may reflect outmigration strategies of fish originating in different parts of the Yukon River system. At the outset of our research, one theory held that the peak of the juvenile Chinook salmon outmigration occurred prior to ice-out on the lower river. Our research suggests that peak migration timing occurs sometime after ice-out and may be related to temperature, prey availability, or other factors.


